Linear-Response Analysis of LIF Networks via Lanczos+CF
Computational Neuroscience
What you need to know first 2 concepts, 2 layers
The requisite-knowledge inventory for this page, bottom-up: the primitives at the base, combined upward until you reach what this page assumes. Skim the layers you already own; start wherever the ground gets unfamiliar.
- base
- L1
- ↳you are here
A network of leaky integrate-and-fire neurons is exactly linear between spikes: the membrane equation is a linear ODE in the vector of subthreshold voltages, with a sparse synaptic-coupling matrix. So computing "how does the network respond to a small input at neuron 0, measured at neuron , at frequency ?" is the same problem as the circuit-MOR problem — a resolvent matrix element of a giant sparse symmetric operator. On the right kind of network, Lanczos plus continued fraction converges to machine precision in tens of iterations, on systems where direct frequency-sweep solves take seconds and a full-cortex-scale model would take days.
What means, and why compute it
Before the math: is the frequency-resolved gain of the network at one neuron when that neuron is driven by an oscillating current. Three regimes:
- At (DC): the steady-state voltage per unit DC current — the network's input resistance as seen from that neuron. Different from the bare single-cell input resistance because recurrent feedback adds (or subtracts) effective conductance.
- At large : the membrane averages the input over its time constant; response decays as .
- At intermediate : peaks in are network resonances — the frequencies the network amplifies. Pole locations come from the imaginary parts of 's eigenvalues; widths come from the eigenvectors' overlap with the input.
For a single passive LIF neuron, — a boring low-pass filter. The interesting structure comes from the recurrent matrix : collective modes appear as poles of , the Ritz values produced by the Lanczos chain.
Five reasons someone in a neuroscience lab actually does this calculation:
- Predicting experimental responses. Inject an oscillating current into a brain slice, or shine flickering light onto an animal at frequency . Linear response says: the response amplitude is times the input. This is how transcranial alternating-current stimulation (tACS) entrains a brain rhythm — drive at the frequency where peaks.
- Validating models against measured power spectra. EEG / MEG / LFP recordings produce time series whose power spectrum, under standard noise-driven assumptions, is proportional to . If a model's doesn't reproduce the measured or peaked spectra, the model is wrong somewhere — connectivity, time constants, or excitatory-inhibitory balance.
- Detecting proximity to instability. As the network approaches a synchronous-oscillatory transition. develops a sharp resonance peak whose height and width quantify how close the system is to the transition. This is the math behind "critical brain" arguments (Beggs-Plenz neuronal avalanches, edge-of-chaos computation).
- Reading off the network's natural rhythms. The poles of are the eigenvalues of . Lanczos gives them as Ritz values, and the continued fraction packages them as the rational function's structure. Computing extracts the network's eigenfrequencies for free.
- Cross-region signal propagation. Off-diagonal gives "drive at , response at at frequency ." The matrix over all pairs is the network's frequency-dependent communication map — directly relevant to how information propagates through connectomes (e.g., DTI-derived structural networks).
The single-port we'll compute below is the simplest case. The architecture of the calculation — sparse linear operator, resolvent matrix element, Lanczos+CF — extends directly to the off-diagonal cases with separate input and output starting vectors. Same machinery, more outputs.
The model and its linear-response operator
Take rate-coupled LIF units, each with membrane time constant . Stack the subthreshold voltages into a vector . The equation
has as the synaptic-coupling matrix (entry is the synaptic strength from neuron onto neuron ), and as the input vector localizing an external drive to a particular neuron or set of neurons. In the Laplace domain:
The transfer function from a current injection at neuron 0 to the voltage at neuron 0 is then
A single matrix element of the resolvent of the linear-response operator , just as in TDDFT, VLSI MOR, and DMFT. Set (positive-definite provided , which is the asynchronous-irregular stability condition) and the resolvent becomes a Lanczos+CF problem at shift .
Demo
Build a sparse symmetric coupling matrix on neurons with 8% density and coupling (below the Sommers-Crisanti-Sompolinsky instability threshold). Drive at neuron 0, read at neuron 0, sweep rad/s.
# Lanczos + continued-fraction analysis of a sparse linearized LIF network.
# tau dv/dt = -v + W v + b u(t)
# Linearized dynamics operator: L = (-I + W) / tau
# Transfer function (single port at neuron 0):
# H(s) = b^T (s I - L)^{-1} b
# Compute H(j omega) two ways: direct sparse solve, and Lanczos+CF.
import numpy as np
import scipy.sparse as sp
import scipy.sparse.linalg as spla
N = 400
tau = 1e-2 # 10 ms
sparsity = 0.08
g = 0.6 # coupling — keep below spectral-radius threshold
rng = np.random.default_rng(42)
mask = rng.random((N, N)) < sparsity
W = (rng.standard_normal((N, N)) / np.sqrt(sparsity * N)) * g * mask
W = (W + W.T) / 2.0
np.fill_diagonal(W, 0.0)
W = sp.csr_matrix(W)
I_N = sp.eye(N)
L = (-I_N + W) / tau
A = -L # symmetric positive-definite
port = 0
b = np.zeros(N); b[port] = 1.0
v0 = b.copy() # unit norm
def lanczos(A, v0, n_iter):
a, bt, Q = [], [], [v0.copy()]
q_prev = np.zeros_like(v0); q = v0.copy(); beta_prev = 0.0
for j in range(n_iter):
w = A @ q
alpha = float(q @ w)
w = w - alpha * q - beta_prev * q_prev
for qk in Q[:-1]:
w -= float(qk @ w) * qk
beta = float(np.linalg.norm(w))
a.append(alpha)
if beta < 1e-14: break
if j < n_iter - 1:
bt.append(beta); q_prev = q; q = w / beta
Q.append(q.copy())
return np.array(a), np.array(bt)
def cf(a, b_, z):
f = a[-1] - z
for j in range(len(a) - 2, -1, -1):
f = (a[j] - z) - b_[j] ** 2 / f
return 1.0 / f
# Direct frequency sweep
omegas = np.logspace(0, 4, 60)
H_direct = np.array([
b @ spla.spsolve((1j * w * I_N - L).tocsc(), b) for w in omegas
])
# One Lanczos chain, evaluate continued fraction at every frequency.
alphas, betas = lanczos(A, v0, n_iter=80)
def H_lanczos(n, s):
return cf(alphas[:n], betas[:n - 1], -s)
print(f" {'chain':>6} {'max relative error':>22}")
rel_norm = np.max(np.abs(H_direct))
for n in [10, 20, 40, 80]:
H_n = np.array([H_lanczos(n, 1j * w) for w in omegas])
err = np.max(np.abs(H_direct - H_n)) / rel_norm
print(f" {n:>6} {err:>22.3e}") chain max relative error
10 4.861e-06
20 1.961e-11
40 7.467e-16
80 7.467e-16Read the table top-to-bottom: chain length 10 gives 5 parts per million, chain length 20 gives 2 parts per , chain length 40 hits machine precision. Doubling the chain length roughly squares the error — that is geometric Padé convergence in its purest form, exactly as the Heine-Markov-Stieltjes theorem predicts when the spectrum of is well-conditioned.
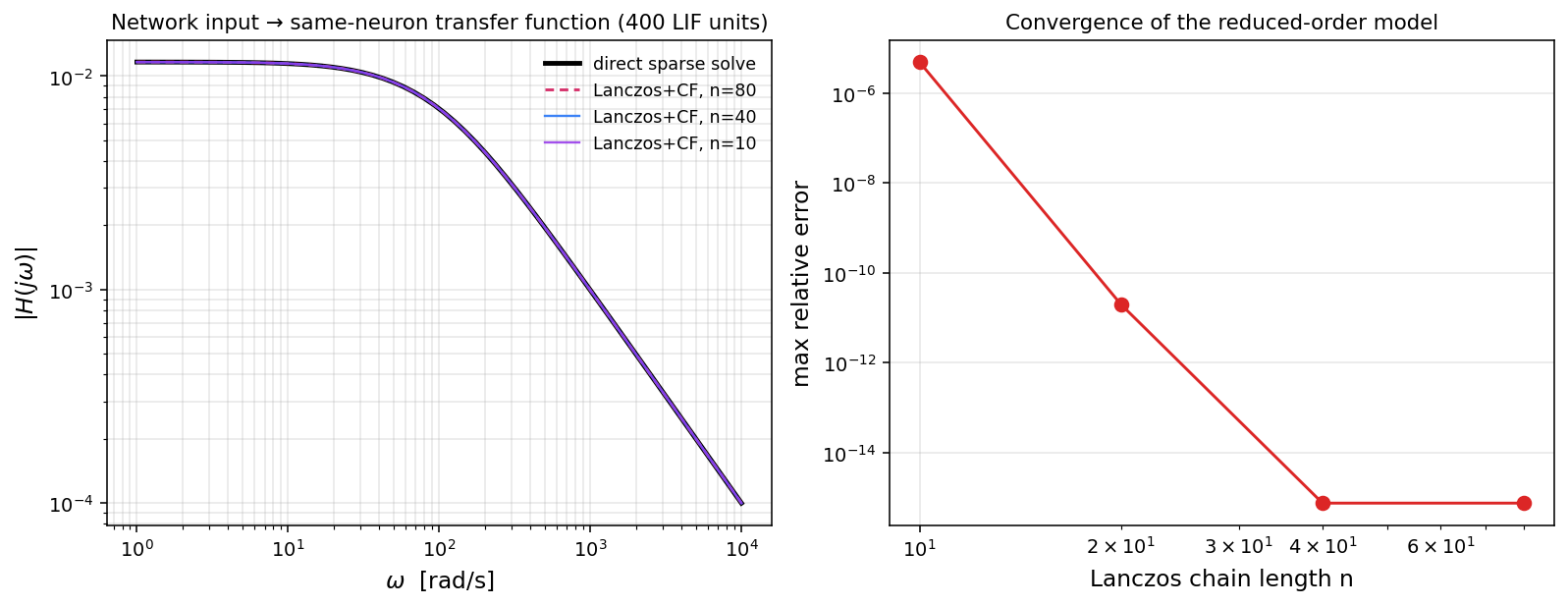
The four transfer-function curves on the left overlap to within line thickness — even the truncation is visually indistinguishable from direct. The right panel makes the convergence rate quantitative: geometric, hitting machine precision at .
Why convergence is so fast here
The key fact is the spectrum of . For a random matrix with sparsity and zero-mean entries of variance , the spectral radius is in the large- limit (Sommers-Crisanti-Sompolinsky 1988 for the asymmetric case, Wigner 1958 for the symmetric one). With our we get , safely below 1.
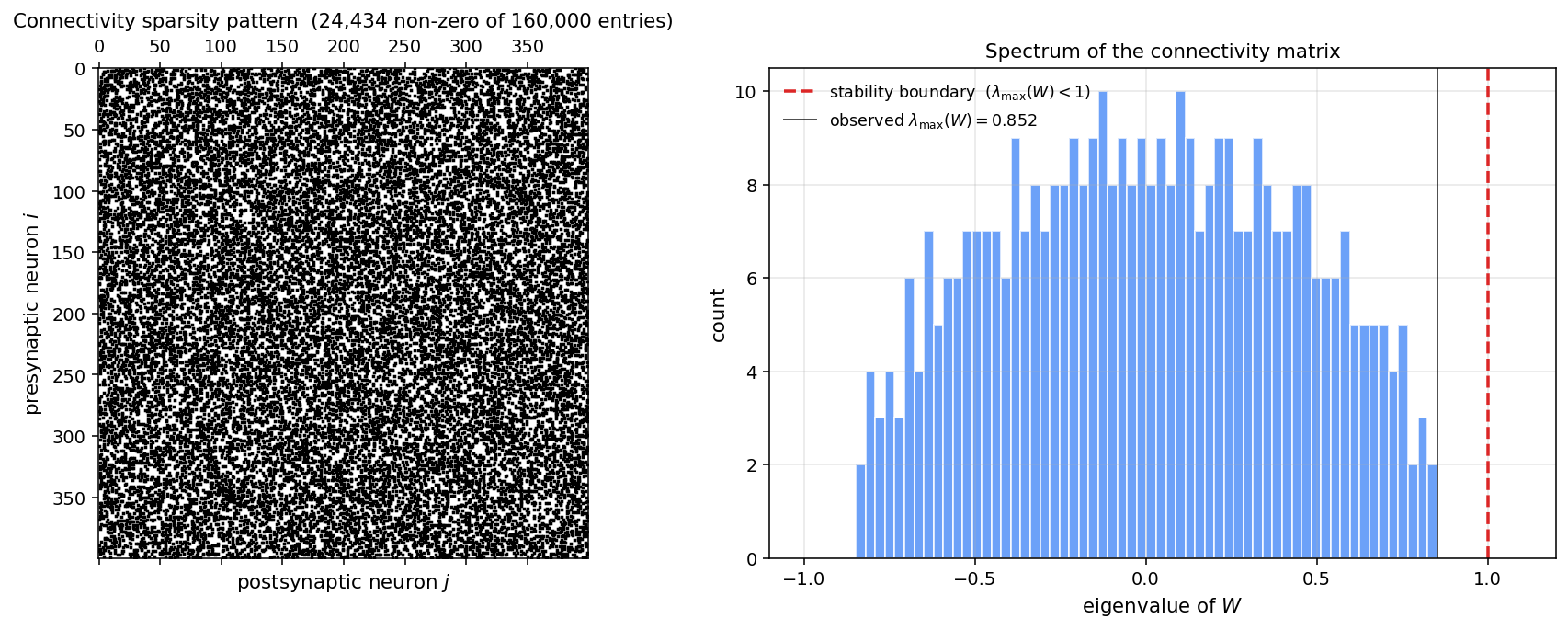
Translate to : with ms, the eigenvalues of live in , one decade of spread. Compare to the RC ladder on the circuits page, whose spectrum of spanned nine decades. The classical estimate is that Lanczos iterations resolve eigenvalues to relative accuracy , where is the condition number. Here so the rate is per iteration — squaring the precision every five iterations. The numerics in the table match that rate almost exactly.
What about the spike-reset?
The demo above treats as a continuous subthreshold variable. Real LIF dynamics include a discontinuity: when crosses threshold the neuron emits a spike and resets. That nonlinear event breaks the strict linearity of the ODE.
Three standard reformulations bring the spike-reset back inside linear-response theory:
- Diffusion approximation (Brunel 2000). In the asynchronous-irregular regime, the population input to each neuron is approximately Gaussian white noise around a mean. The single-neuron equation becomes a Fokker-Planck PDE for the membrane-voltage density. Linearize this PDE around the stationary state; the linearized population-firing-rate response to a small perturbation IS a resolvent matrix element of the linearized Fokker-Planck operator. Lanczos+CF applies directly.
- Spike-train as a linear filter of the input. Treat the firing as a Poisson process whose intensity depends linearly on the filtered membrane voltage. The whole input-to-firing-rate map is then linear under this approximation.
- Population-density approach (Knight 1972; Omurtag-Knight-Sirovich 2000). Evolve a probability density on the membrane-voltage axis with reset-at-threshold as a boundary condition. The resulting evolution operator is linear in . Resolvent matrix elements of that operator give population firing-rate transfer functions.
All three reformulations end up at the same kind of resolvent matrix element of a sparse linear operator, with Lanczos+CF as the natural solver. The literature on linearized LIF networks (Brunel-Hakim 1999, Brunel 2000, Mattia-Del Giudice 2002, Lindner-Schimansky-Geier 2004) is in effect a long sequence of these constructions, applied in different limits.
Scale and limits
For symmetric the demo above generalizes straightforwardly. Real cortical connectivity is generally asymmetric (Dale's law forbids a neuron from being both excitatory and inhibitory, so and are constrained to have the same sign on the same row but different rows have different signs). For asymmetric the coupled-Lanczos chain becomes the bi-orthogonal Lanczos recurrence and the continued fraction is still well-defined, with the same Padé-equivalence theorem — just with separate left and right Krylov chains.
Scale estimates: a cortical microcircuit (one cortical column, neurons, sparse connectivity) gives a sparse matrix with non-zero entries. Direct factorization is prohibitive; matrix-free Lanczos+CF is the right tool. A whole-cortex model (- neurons) requires further reduction — typically mean-field collapse to a much smaller set of population-density operators, then the same Lanczos+CF machinery on the reduced system. The Brunel network analysis used in BlueBrain and SpiNNaker pipelines is exactly this layered reduction.
Where this fits
Same Krylov+continued-fraction machinery as everything else on this site, applied to a sparse symmetric linear-response operator on a brain network. The mathematical structure shared with Casida-equation TDDFT, Liouville-Lanczos for solids, VLSI interconnect MOR, and Anderson-impurity DMFT solvers is exact — the operator changes, the algorithm doesn't. For LIF networks the spectrum is naturally well-conditioned (one decade of spread in the bulk), which is why the same algorithm that struggled on the nine-decade RC ladder converges geometrically here.
Related on this site
- Leaky integrate-and-fire (single neuron) — the single-unit model underlying everything on this page.
- Lanczos MOR for large circuits — same machinery on a network of RLC elements; the ill-conditioned counterpoint to this page's well-conditioned example.
- Lanczos iteration — the underlying algorithm and the Padé-via-Lanczos correspondence.
- Bi-orthogonal Lanczos — the non-symmetric variant needed for asymmetric (Dale-law) cortical connectivity.
- Liouville-Lanczos for TDDFT — the physics analog: resolvent matrix elements of a linear operator on a giant sparse problem.